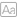2023年5月25日德国马克斯·普朗克生物智能研究所Rüdiger Klein团队在ScienceAdvance发表了“Transcriptomics reveals amygdala neuron regulation byfasting and ghrelin thereby promoting feeding”,该研究在转录组学揭示了杏仁核神经元通过禁食和胃饥饿素调节从而促进摄食行为,CeA神经元和微环路的功能障碍可能是病理进食行为的基础。
 (相关资料图)
(相关资料图)
中央杏仁核(CeA)可进一步分为内侧中央核(CeM)、中央囊区(CeC)和外侧中央核(CeL),CeA由许多抑制性神经元组成,这些神经元控制防御和食欲行为,包括进食。但是细胞类型的转录组特征及其与功能的联系知之甚少,在本研究中利用单核RNA测序描述了9个CeA细胞簇,其中4个与食欲行为有关,2个与厌恶行为有关。为了分析食欲性CeA神经元的激活机制,对表达5-羟色胺受体2a (Htr2a)的神经元(CeAHtr2a)进行了表征,这些神经元包括三个食欲簇,并且先前被证明可以促进进食。体内钙成像显示CeAHtr2a神经元在禁食、胃饥饿素激素和食物的存在下被激活。此外,这些神经元是胃饥饿素的摄氧作用所必需的。食欲性CeA神经元响应禁食和胃饥饿素投射到臂旁核(PBN),引起目标PBN神经元的抑制。这些结果说明了CeA神经元的转录组多样化与禁食和激素调节的摄食行为相关。
图一 小鼠中央杏仁核的转录组细胞类型分类
作者首先使用单核RNA测序进行鉴定CeA区域中亚型的多样性。从8周龄小鼠中获得两个数据集。第三个数据集来自Htr2a-Cre转基因小鼠,以确定哪些亚型包括在CeAHtr2a群体中。作者获得了3325个单核转录组,并利用主成分分析(PCA)对标度基因表达的影响数据,一共获得了16个集群,并根据多个标记基因的共表达将它们划分为不同细胞类别。GABA能神经元是主要的细胞类型,形成11个簇,一小部分神经元表达兴奋性标记物Scl17a7。重新聚集了2488个GABA能神经元,确定了不属于CeA的簇。对于属于CeA的其余9个簇,基于高可变基因在细胞类型中的表达相关性构建了一个分类树。第一次划分将Prkcd、Cdh9/Calcrl和Dlk1神经元从其他簇中分离出来,即Prkcd、Cdh9/Calcrl和Dlk1神经元先前被证明存在于CeL中,主要参与厌食和其他厌恶功能,但也参与食欲/情感行为。第二次分类从剩余的四个细胞簇中分离出表达NTS/Tac2和sst的细胞。许多NTS/Tac2细胞低水平表达Sst,但Sst表达最高的细胞形成一个单独的簇。大多数集群是在CeM中发现的,CeM是CeA的一个分区域。同时证实CeAPnoc和CeAHtr2a神经元部分重叠,54%的CeAHtr2a神经元是伤害肽阳性,34%的伤害肽阳性神经元属于CeAHtr2a群体。
图二 禁食或胃饥饿素激活CeA-Htr2a神经元
作者通过单细胞测序技术发现CeA脑区不同亚区存在9种抑制性神经元,CeC-Calcrl和CeL-PKCδ主要与厌恶行为相关,CeL-NTS、CeL-Tac2、CeL-Sst、CeM-Il1Rapl2神经元主要与食欲相关,CeA-Htr2a神经元与摄食行为相关。免疫荧光实验发现在禁食后小鼠CeA神经元活性增强。腹腔注射胃饥饿素促进小鼠摄食,也会引起饱腹小鼠CeA神经元激活增多。进一步实验发现胃饥饿素主要激活CeA-Htr2a神经元,几乎不激活CeA-PKCδ神经元。光纤钙成像技术发现在进食过程中CeA-Htr2a神经元活性增强。在无食物供应情况下,胃饥饿素也能增加CeA-Htr2a神经元活性,在给与食物供应后,这种活性的增强更加明显。
图三 胃饥饿素刺激表达5-羟色胺受体2a的神经元
作者接下来利用脑切片电生理记录研究了食欲性CeA神经元的激活机制。结果显示,在禁食动物中,CeAHtr2a神经元的兴奋性与喂食动物相比显著增加,禁食动物的放电频率高于喂食动物。为了研究动作电位频率的差异是否可能由内源性胃饥饿素水平升高引起,作者对灌注胃饥饿素3分钟的脑切片进行了记录。结果表明,胃饥饿素诱导CeAHtr2a神经元放电,在胃饥饿素应用几分钟后,大多数神经元去极化。胃饥饿素增加了兴奋性神经传递的频率,但不影响振幅变化。相反,没有观察到胃饥饿素诱导的CeAPKCδ神经元兴奋性神经传递的变化。这些结果表明,胃饥饿素诱导谷氨酸从突触前输入特异性地释放到CeAHtr2a神经元,从而增加这些食欲神经元的兴奋性。
图四 CeA-Htr2a神经元促进摄食
电生理实验结果表明:与进食小鼠相比,禁食小鼠CeA-Htr2a神经元放电活动增强,孵育胃饥饿素也能明显增加CeA-Htr2a神经元放电活动,增强自发性兴奋性突触后电流频率,但几乎不影响CeA-PKCδ神经元电活动。化学遗传学慢性抑制CeA-Htr2a神经元活性后并不影响小鼠在饱腹状态下的摄食行为,但可完全阻断胃饥饿素引起的促摄食行为效应。在激活CeA-Htr2a神经元可进一步增强饥饿素引起的促摄食行为效应,这种增强作用可被饥饿素拮抗剂所阻断。这些结果表明CeAHtr2a神经元的活动以双向方式影响外源性胃饥饿素诱导的食物摄入。
图五 禁食和胃饥饿素激活CeA→PBN投射,抑制PBN神经元
为了特异标记投射到PBN上的CeAHtr2a神经元,作者将将逆行病毒注射到Htr2a-Cre小鼠的PBN中,CeAHtr2a神经元在给予胃饥饿素后sEPSC的频率增加。如果PBN受到CeAHtr2a神经元的抑制控制,则PBN神经元应受到GABA释放的抑制。作者发现禁食后PBN神经元的动作电位频率降低。脑切片中给予胃饥饿素发现大部分PBN神经元IPSC频率增加,禁食对PBN神经元的抑制作用与胃饥饿素相似。与喂食动物相比,禁食动物的IPSC频率增加。这些结果表明CeAHtr2a→PBN投射被禁食和外源性胃饥饿素激活,导致其靶向PBN神经元受到抑制。CeAHtr2a→PBN投射可能与下丘脑弓状区的GABA能刺鼠关联蛋白(AGRP)神经元平行作用,后者也投射到PBN并通过抑制其靶向PBN神经元来促进进食。
图六 胃饥饿素通过CeA-Htr2a→PBN环路促进摄食
已有研究表明CeA-Htr2a神经元投射到臂旁核PBN。作者发现PBN的轴突中FosCh的表达增加,表明禁食激活了CeA→PBN投射,逆行示踪实验发现PBN逆行投射到CeA的神经元在孵育胃饥饿素后可增强兴奋性突触传递作用。Htr2a-cre工具小鼠中也发现胃饥饿素或禁食也能激活CeA-Htr2a→PBN环路。此外,禁食状态下或孵育胃饥饿素后PBN神经元放电活动减弱。化学遗传学慢性抑制CeA-Htr2a→PBN环路后可明显阻断胃饥饿素的促摄食作用。总之,这些结果表明禁食激活CeA→PBN投射,从而抑制其靶向PBN神经元。该结果进一步表明,CeA→PBN投射的活动对饥饿素诱导的食物摄入有重要作用。
图七 脑状态相关差异基因表达的鉴定
为了剖析更细微的转录组变化对禁食的反应,作者迭代地聚集了CeA的GABA能神经元。CeLNTS/Tac2簇在禁食后表现出强烈的细胞状态变化,表明这种细胞类型的基因表达发生了明显改变。为了揭示禁食动物与自由喂养动物之间细胞类型特异性的变化,作者进行了差异基因表达分析,发现所有群集在禁食状态下差异表达基因数量均有增加。然而,到目前为止,CeLNTS/Tac2集群在上调和下调基因中显示的差异表达基因数量最多,其次是CeLPKCδ。对CeLNTS/Tac2和CeLPKCδ群体中差异表达基因的富集分析揭示了与神经元活性包括突触、离子转运、激酶活性和肌动蛋白结合和转录因子相关。作者接下来评估了禁食是否促进CeAHtr2a神经元树突棘的形成,结果发现,禁食小鼠的CeAHtr2a神经元的树突棘密度比喂食小鼠增加了约26%。总之,这些结果表明,CeA神经元的一个子集通过动态基因表达变化和树突棘数量的增加对禁食行为做出反应。
综上所述,该研究在转录组分类学说明了CeA神经元的多样化,并显示了CeA中食欲神经元的活动如何受到内感受性线索(禁食和胃饥饿素)和食物的存在的调节。食欲性CeAHtr2a神经元对胃饥饿素的促食作用至关重要,它们投射到PBN上,引起目标PBN神经元的抑制。研究结果揭示了CeA在胃饥饿素系统调节摄食行为中的作用。此外,该结果为进一步研究特定的CeA神经元群如何与享乐和稳态进食回路相互作用以及情绪状态-食物摄入和奖励之间的相互作用奠定了基础。以往杏仁核的研究主要在调控抑郁、焦虑、恐惧记忆等领域,本文揭示了杏仁核通过胃饥饿素信号调控摄食行为的新机制。
文章来源
/doi/10.1126/sciadv.adf6521
关键词: